Erfahrungen (und Medikamente) verändern nicht nur den Hirnstoffwechsel, sondern auch die Hirnsubstanz.
In diesem Artikel erfahren Sie wie die Übertragung von Signalen in unserem Nervensystem grundlegend funktioniert, wie Lernprozesse das Gehirn und die Synapsen beeinflussen und dauerhaft verändern und wie Psychopharmaka und andere Medikamente an den Synapsen wirken.
Für viele dürfte überraschend sein, dass medikamentöse Therapien, etwa mit Psychopharmaka und nicht-medikamentöse Therapien wie die Psychotherapie, ähnlichen Angriffspunkt im Gehirn haben. Die Kenntnis der Vorgänge an den Nervenzellen bringt Biologie und Psychotherapie einander näher. Lernen und Erfahrung verändert das Gehirn genau auf ähnliche Art und Weise wie bestimmte Medikamente.
Durch die Veränderbarkeit (Plastizität) des Gehirns bestimmen Gene nur bedingt unser Schicksal. Der alleinige Glaube an die alles bestimmenden Gene ist ebenso überholt, wie die Leugnung, dass sie wesentlichen Einfluss haben. Informationen aus der Umgebung verändern die Hirnsubstanz und die Nutzung der genetischen Information. Dadurch wird auch das Verhalten verändert. Reiz und Reaktion sind bei Menschen kein einfacher Reflex, sondern ein kompliziertes, bisher nur teilweise verstandenes Gefüge von komplizierten biologischen Vorgängen.
Gefühle und Denken haben auch eine biochemische Grundlage. Das Verstehen dieser Grundlagen hilft auch psychosoziale Zusammenhänge zu verstehen und an deren Verbesserung zu arbeiten. Komplexe Verhaltensweisen von Menschen haben viele Erklärungen auf unterschiedlichsten Ebenen. Biologie, Biochemie, Genetik, Psychologie, Soziologie und Psychiatrie sind keine Gegensätze, sondern stellen verschiedene Aspekte in der Erklärung menschlichen Verhaltens dar.
Die Synapse
Der Begriff Synapse wurde 1897 von Sherrington für die Verbindungsstelle zwischen 2 Nervenzellen eingeführt. An dieser werden Informationen zwischen den Zellen ausgetauscht.
Ein Neuron oder eine Nervenzelle besteht aus einem Zellkörper (Perikaryon, oder Soma) mit Zellkern dem (Nucleus) und einem langen Hauptfortsatz, dem Axon. Das Axon kann einige Millimeter und bis zu einem Meter lang werden. Vom Zellkörper gehen viele kurze Fortsätze aus, die Dendriten genannt werden und an denen andere Neuronen mit ihrem Axon „ankoppeln“ können. Die Dendriten vergrößern dabei die Oberfläche des Neuron und bilden zusammen mit dem Soma (Zellkörper) den Ort des Erregungsempfangs eines Neurons. Sender von der Zelle ist die axonale Nervenendigung.
Damit ein Neuron eine Information an ein anderes Neuron weiterleiten kann, besitzt jedes Axon an seinem Ende zahlreiche Verästelungen, an denen sogenannte Endknöpfchen sitzen. Diese liegen an der Oberfläche anderer Nerven oder Muskelzellen beinahe auf und bilden so die sog. Synapse. Der Spalt oder Zwischenraum an diesen Verbindungsstellen zwischen zwei Nervenzellen ist etwa 20-30 nm breit und wird Synapsenspalt oder synaptischer Spalt genannt.
Über den synaptischen Spalt wird das zu übertragende Signal mit Hilfe verschiedener Botenstoffe, so genannter Transmitter oder Neurotransmitter geleitet („Transmitter“= Überträger, im Nervensystem „Neurotransmitter“). Dieser Spalt enthält transmitterabbauende Enzyme. Nach Übertragung des Signals wird der Transmitter entweder durch diese Enzyme abgenaut oder wieder in den Axon aufgenommen („Reuptake“).
Synapsen kommen als Verbindungsstelle zwischen 2 Nervenzellen, zwischen Nervenzellen und Muskelzellen oder Nervenzellen und Sinnenszellen vor. Sie bilden die (elektronen-) mikroskopisch kleine Grundlage menschlicher Lernvorgänge, sind die kleinen Schaltstellen unseres Bewusstseins wie jeder Wahrnehmung, Interpretation, Kommunikation oder Bewegung.
Bei den gut erforschten chemischen Synapsen wird als Folge der elektrischen Erregung eines Neurons an dessen Synapse eine chemische Substanz freigesetzt. Diese Transmitterausschüttung erfolgt in rasanter Geschwindigkeit von 1/5000 Sek. Der erste Transmitter, der genauer bekannt wurde, war Acteylcholin. An seinen Beispiel werden Transmitter noch immer in Biologiebüchern näher erklärt. Zwischenzeitlich wurden jedoch mehr als 100 Neurotransmitter identifiziert, vermutlich gibt es mehrere tausend.
Funktionsweise der Signalübertragung über Transmitter
Neurotransmitter werden in der Zelle (unter Beteiligung des endoplasmatischen Reticulums und des Golgi-Apparats) synthetisiert und in kleinen „Membranbläschen“ (sog. Vesikel) gespeichert. Sie müssen darin oft weite Strecken vom Zellkörper zur Synapse transportiert werden. Ein ankommendes Aktionspotential (elektrischer Impuls) erregt die Membran im Bereich des Axon- oder Dentritenendköpfchens und aktiviert dadurch Kalziumkanäle (Ca2+ – Kanäle), die einen Einstrom von Ca2+ – Ionen aus der umgebenden Zellflüssigkeit ermöglichen.
Die erhöhte Ca2+-Konzentration löst die Wanderung der Vesikel an die präsynaptische Membran und die Ausschüttung des Transmitters aus („Exocytose der Vesikel“). Die Vesikel verschmelzen dabei mit der präsynaptischen Membran (fusionieren mit der Zellmembran) und und öffnen sich dabei, um den Neurotransmitter in den Spalt freizugeben.
Diese Neurotransmitter wiederum bewirken, dass es in dem über die Synapse verbundenen Neuron ebenfalls zu einer elektrischen Erregung kommt. Die Synapse und die Neurotransmitter dienen damit der chemischen Übertragung der fortgeleiteten elektrischen Aktivität von einer Nervenzelle auf die nächste (von der präsynaptischen Nervenzelle (prae=vor) auf die postsynaptische Zelle (post= hinter).
An einer Synapse kann die Erregung nur in eine Richtung übertragen werden. Synapsen haben damit eine Art Ventilwirkung. Im Endknopf der Synapse befinden sich Mikrotubuli, die im Zytoplasma synthetisierte Neurotransmitter in den Endknopf transportieren und Vesikel, in denen die Neurotransmitter gespeichert werden. (s.u.)
Wirkung der Neurotransmitter am Rezeptor
Die Wirkung eines Neurotransmitters (exzitatorisch = erregend oder inhibitorisch = hemmend) an der postsynaptischen Zelle hängt nicht von den chemischen Eigenschaften des Transmitters ab, sondern von den Eigenschaften des Rezeptors. Der wichtigste Bestandteil der postsynaptischen Membran sind transmitterspezifische Rezeptoren. Es gibt aber durchaus überwiegend erregende (z.B. Glutamat u. Acetylcholin) oder überwiegend hemmende (GABA, Glycin) Neurotransmitter.
Rezeptoren bilden mit dem Überträgerstoff einen funktionalen Komplex. Agonisten sind Substanzen, die stimulierend auf einen Rezeptor wirken, Antagonisten sind Substanzen, die hemmend auf einen Rezeptor wirken. Rezeptoren besitzen eine bestimmte Selektivität und Affinität. Substanzen, die zu einem bestimmten Rezeptor eine hohe Affinität zeigen, werden als Liganden bezeichnet.
An den Dendriten und den Zelleibern der meisten Nervenzellen findet sich ein Gemisch aus inhibitorischen (hemmenden) und excitatorischen (erregenden) Synapsen. Der jeweilige Erregungszustand solcher Nervenzellen stellt demnach eine Integration der aus unterschiedlichen Richtungen eingetroffenen Informationen dar.
Rezeptoren werden in ionotrope Rezeptoren und metabotropen Rezeptoren unterschieden.
Ionotrope Rezeptoren
Die ionotropen Rezeptoren können nachdem ein spezifischer Transmitter gebunden hat direkt ein elektrisches Potential aufzubauen, da sie strukturell zugleich einem Ionenkanal entsprechen. Ionenkanäle sind hochspezialisierte Öffnungen, durch die bei Bedarf Ionen strömen können. Sie sind spezifisch eine ganz bestimmte Ionenart (Natrium-, Kalzium- und Kaliumkanäle). Bei einer elektrischen Erregung einer Nervenzelle kommt es zu einer Öffnung der unterschiedlichen Ionenkanäle in einem genau festgelegten zeitlichen Ablauf, beginnend mit Natriumkanälen, gefolgt von Kaliumkanälen.
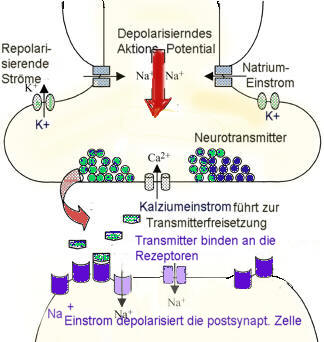
Die in der Ausgangssituation bestehenden Konzentrationsunterschiede der verschiedenen Ionenarten zwischen Zellinnerem und Extrazellularraum führen dabei zu raschen Ionenverschiebungen entsprechend den Konzentrationsgefällen. Natrium fließt nach innen, Kalium nach außen. Nach Beginn des Natriumstroms potenziert sich dieser schnell. Die Natriumkanäle haben dabei eine festgelegte Öffnungszeit von 1-2 ms. Danach schließen sie und bleiben für eine Refraktärzeit geschlossen.
Wenn durch die geöffneten Natriumkanäle große Mengen Natrium nach außen geflossen ist, kommt es zu einer Depolarisierung, da die Kaliumkanäle zunächst nicht geöffnet sind. Erst am Maximum des Natriumflusses öffnen die Kaliumkanäle ohne Zeitlimit um das Konzentrationsgefälle wieder auszugleichen. Durch Ausstrom von Kaliumionen kommt es zu einer vorübergehenden Hyperpolarisierung der Membran.
Im Ruhezustand ist also die postsynaptische Nervenzelle (hinter der postsynaptischen Membran) negativ geladen, dies auch, da es dort mehr negative Protein- Ionen (Anionen) gibt als positive Kalium Ionen. -> Es liegt also eine Spannung von -30 bis -100 mV vor, die als Ruhepotential bezeichnet wird. Die postsynaptische Membran entspricht damit dem Di-elektrikum beim Kondensator.
Wird die Synapse erregt, werden vom Endknopf Neurotransmitter über den synaptischen Spalt zur postsynaptischen Nervenzelle geschickt. Dadurch wird deren Membran kurzzeitig durchlässig (permeabel) für die positiven Natrium-Ionen, die dann schnell aus dem synaptischen Spalt in die Nervenzelle einströmen. Das Membranpotential wird durch den Stromfluss für kurze Zeit „aufgehoben“ bzw. auf 0 V gebracht oder bis 30mV hyperpolarisiert. Es entsteht ein so genanntes Aktionspotential, die Erregung wird nun in der Zielnervenzellen wieder auf elektrisch fortgeleitet. .
Die Ladung der durch die Ionenkanäle wandernden Ionen bestimmt, ob es sich bei der Synapse um eine erregende oder hemmende Synapse handelt. Positiv geladene Ionen erhöhen das Potential im Inneren der Zelle und damit die Wahrscheinlichkeit, dass ein Aktionspotential ausgelöst wird. In diesem Fall nennt man die Synapse erregend oder excitatorisch. Bei einer hemmenden oder inhibitorischen Synapse wird das Potential im Inneren der Zelle durch negativ geladene Ionen erniedrigt und die Bereitschaft zu feuern wird herabgesetzt.
Ob ein Transmitter die postsynaptische Membran depolarisiert oder hyperpolarisiert hängt vom Rezeptortyp ab. Je nach Ionenkanal, mit dem der Rezeptor gekoppelt ist, öffnet der Transmitter Natriumkanäle (–> Depolarisierung, excitatorische Wirkung) oder Kalium- bzw. Chlorkanäle (–> Hyperpolarisierung, inhibitorische Wirkung).
Ionenkanalkrankheiten (Channelopathies) sind Erkrankungen, bei denen diese Ionenkanäle und damit auch die Signalübertragungen an den Synpasen nicht richtig funktionieren.
Metabotrope Rezeptoren
Metabotrope Rezeptoren können nur indirekt ein Potential aufbauen nachdem ein spezifischer Transmitter gebunden hat. Dies funktioniert über die Zwischenschaltung einer „second messenger“-Kaskade, die z. B. G-Proteine, Adenylatzyklase, cAMP und cAMP-abhängige Kinasen, die dann Kanalproteine phosphorylieren, umfassen.
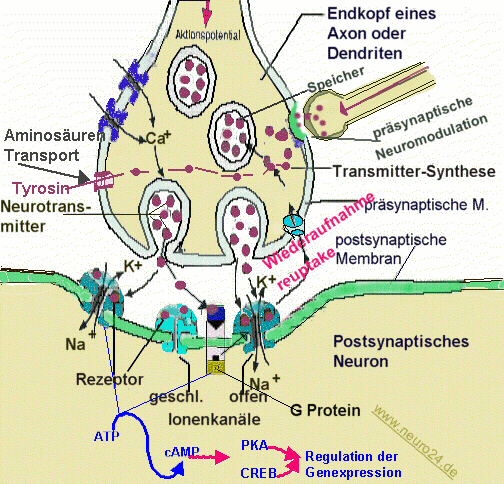
Im ZNS wirken viele Transmitter wie Dopamin und Noradrenalin indirekt, indem sie die Konzentration eines Second messenger erhöhen oder senken, der dann seinerseits die elektrischen oder biochemischen Wirkungen auslöst. Dieser Second messenger ist in vielen Fällen das zyklische Adenosinmonophosphat ( cyclo-AMP ). Der Neurotransmitter dockt dabei an das Rezeptorprotein an, welches daraufhin ein G-Protein auf der Innenseite der Membran aktiviert. Das G-Protein seinerseits aktiviert ein Enzym (Adenylatcyclase), welches aus ATP den sekundären Botenstoff cAMP synthetisiert. Das cAMP setzt sich in das so genannte allosterische Zentrum des Ionenkanals, der sich daraufhin öffnet.
Die Konzentration von cAMP kann von Neurotransmittern z.B. von Dopamin, Serotonin, Muscarin; Acetylcholin und Noradrenalin verändert werden kann, da deren Rezeptoren in der postsynaptischen Membran an das Enzym Adenylatcyclase gekoppelt sind. Die Adenylat-cyclase ist inaktiv, bis der entsprechende Transmitter sich an den Rezeptor bindet. Da ein Molekül Adenylatcyclase etwa tausend Moleküle cyclo- AMP katalysiert, wird das schwache Signale der Transmitter-Rezeptor-Reaktion deutlich verstärkt.
Nicht nur Neurotransmitter auch Hormone, Neuromodulatoren und Wachstumsfaktoren wirken über cAMP.
cAMP und DARPP-32
cAMP ist bei jedem Lernvorgang und jeder Anpassung an unsere Umgebung beteiligt. Eine wichtige Rolle spielt dabei DARPP-32 (Dopamine- and cyclic AMP-regulated phosphoprotein). Es konnte gezeigt werden, dass DARPP-32 eine wichtige Rolle in der Kontrolle von Rezeptoren, Ionenkanälen und andere physiologischen Faktoren spielt, einschließlich der Antwort des Gehirns auf Drogen wie Kokain, Opiate, Koffein und Nikotin. DARPP-32 wird reziprok von Dopamin und Glutamt reguliert.
Dopamin aktiviert DARPP-32 über D1-Rezeptoren und inhibiert es über D2-Rezeptoren. Glutamat inaktiviert DARPP-32 über seinen N-Methyl-D-Aspartate Rezeptor. Bei Patienten mit einer Schizophrenie scheint eine Verminderung von DARPP-32- Proteinspiegeln in den dorsolateralen präfrontalen Rindenregionen eine Rolle in der Krankheitsentstehung zu spielen. Die Hirnregion ist für das abstrakte Denken und das Gefühlserleben wichtig. Nach neueren Hypothesen spielt DARPP-32 auch bei Depressionen eine wichtige Rolle.
Koffein scheintebenfalls über die Phosphorylisation von DARPP-32 zu wirken. Das Protein ist in den Basalganglien besonders konzentriert vorhanden. Dort kommt anscheinend die akitivierende Wirkung des Koffeins zustande.
Second messenger und Genexpression
cAMP führt neben den oben genannten Wirkungen auch zur erhöhten Aktivierung von Proteinkinase A (PKA) und des Transkriptionsfaktors CREB (cAMPresponse-element-binding protein). Transkriptionsfaktoren binden dann an wichtige regulatorische Einheiten von Genen, und beeinflussen wiederum deren Expression in bestimmten Hirnregionen. Second messenger regulieren also auch die Genexpression und verändern damit dauerhaft die Nutzung der Erbsubstanzen der Zellen.
Es ist inzwischen bekannt, dass Transmitter, die über second messenger arbeiten, die Regulationsprotenine für die Transskription phosphorylieren und so die Genexpression verändern.
Die Second messenger induzieren nach dem oben besprochenen nicht nur die Synthese der bereits in der Zelle vorhandenen Proteine sondern auch neuer bisher dort nicht vorhandener Eiweiße, sie nehmen Einfluss auf die RNA. Ein Vorgang der manchmal Tage beansprucht. Hieraus können Veränderungen im Nervenwachstum und eine Veränderung der Synapsen resultieren. Dieser Mechanismus spielt wahrscheinlich beim Langzeitgedächtnis eine große Rolle.
Wirkung von Stickstoffmonoxid (NO) auf die Synapsen
Die Aktivierung von Synapsen führt zur Freisetzung kleiner Mengen des an sich giftigen Gases NO (Stickstoffmonoxid). Dieses wiederum sorgt dafür, dass bei einem ähnlichen Reiz in kurzer Folge diese und ähnliche Synapsen in der Umgebung schneller reagieren und beschleunigen und verbessern damit die Reaktion auf einen Umweltreiz.
NO ist vermutlich an akuten und chronischen entzündlichen und neurodegenerativen Prozessen nützlich wie schädlich beteiligt. Die deletären Folgen eines Schlaganfalls oder einer mechanischen Traumatisierung z. B. werden mit einer Störung der NO-Homöostase in Verbindung gebracht.
Auch primär degenerative Krankheiten, wie die Parkinsonsche und Alzheimersche Krankheit oder die amyotrophe Lateralsklerose, selbst die Migräne und die Entstehung einer Sucht scheinen mit Störungen im NO- Stoffwechsel zusammenzuhängen.
Die molekularen Mechanismen der NO-Wirkung sind außerordentlich komplex. NO kann als Gas nicht in Vesikeln gespeichert werden. Als gasförmiges Radikal vermag NO unter Bindung an Hämgruppen, an Eisen-Schwefel-Cluster und Thiolgruppen mit einer großen Vielzahl von Biomolekülen zu reagieren und diese in ihrer Konstitution, mithin in ihrer Funktionsweise zu verändern.
Außerdem ist NO vielfältig in den Metabolismus reaktiver Sauerstoff- und weiterer Stickstoffspezies eingebunden. Aus NO und dem Superoxidradikal entsteht hochreaktives Peroxynitrit, das in verschiedene, ebenfalls sehr reaktionsfreudige Folgeprodukte umgesetzt werden kann und in mancher Hinsicht sogar als der eigentliche Mediator von NO-Wirkungen verstanden wird.
Arten von Synapsen
Man unterscheidet:
- Neuromuskuläre Synapsen, darunter versteht man die Kontaktstelle eines Motoneurons mit einer Muskelfaser, Überträgerstoff dort ist Acetylcholin. Neuromuskuläre Synapsen sind die Schaltstellen zwischen Nervensystem und Muskel, die jede Art der Bewegung inklusive der Atmung kontrollieren.
- Neuro-neuronale Synapsen zwischen zwei Nervenzellen.
- Je nach den miteinander verbundenen Abschnitten der beteiligten Nervenzellen bzw. Nervenzellfortsätze unterscheidet man bei den Synapsen zwischen Nervenzellen:
- Axo-dendritische Synapsen zwischen einem reizweiterleitenden Fortsatz einer Nervenzelle (Axon) und einem reizempfangenden Fortsatz (Dendrit) einer anderen Nervenzelle.
- Axo-somatische Synapsen zwischen dem Axon einer Nervenzelle und dem Zelleib (Perikaryon = Soma) der nachgeschalteten Nervenzelle.
- Axo-axonale Synapsen zwischen 2 Axonen, dabei bildet ein langes, vorgeschaltetes Axon meist kurz nach dem Abgang des nachgeschalteten Axons vom Perikaryon eine Synapse an diesem aus.
- Somatosomatische Synapsen zwischen den Nervenzellkörpern zweier direkt nebeneinander liegender Nervenzellen.
- Dendrodendritische Synapsen zwischen Dendriten zweier verschiedener Nervenzellen (selten).
- En passant Synapsen: An einem geradlinig verlaufenden Axon wird nach seitlich hin eine Synapse zu einer benachbarten Zelle oder einem anderen Axon gebildet.
- Reziproke Synapsen: Synaptische Endkolben an Axonkollateralen einer Nervenzelle enden am Perikaryon oder an Dendriten derselben Nervenzelle.
Sonderform: Elektrische Synapsen
Eine Sonderform sind die elektrischen Synapsen. Bei einer elektrischen Synapse sind zwei Neuronen direkt durch Kanäle verbunden, über die sich die elektrische Erregung von einem auf das andere Neuron fortpflanzen kann. Das anatomische Korrelat zu elektrischen Synapsen sind die sogenannten Gap-Junctions. Prä- und postsynaptische Membranen sind dicht aneinander gelagert. Es gibt keinen synaptischen Spalt. In beiden Membranen lassen sich elektronenmikroskopisch dicht gepackte Partikel nachweisen (Connexone), die aus je 6 Connexin-Untereinheiten bestehen. Gap junctions sind Ansammlungen interzellularer Kanäle die von Connexinen gebildet werden, eine multigene Familie mit 20 verschiedenen Varianten bei Menschen.
Obwohl elektrische Synapsen im Gegensatz zu chemischen Synapsen im Gehirn weniger verbreitet sind, haben Studien gezeigt, dass insbesondere Interneurone im Hippocampus und Neocortex mittels elektrischer Synapsen kommunizieren (Bruzzone et al. 2003).
Veränderungen der Synapsen bei Lernvorgängen
Unser Gehirn besteht aus etwa 100 Milliarden Nervenzellen. Jede Nervenzelle im Gehirn hat durchschnittlich etwa 10 000 Synapsen mit anderen Nervenzellen. Insgesamt geht man von einer Billiarde Synapsen (1.000.000.000.000.000 Synapsen) im Gehirn aus. Die Zahl der Synapsen nimmt bei Menschen in den ersten 3-6 Lebensjahren zu um dann bis zum jungen Erwachsenenalter wieder abzunehmen. Dies geht allerdings nicht mit einer Verminderung der Lernfähigkeit einher.
Man geht davon aus, dass die Informationsspeicher des Gehirns überwiegend an den Synapse sitzen, dort wo Nervenzellen kommunizieren. Veränderungen an den Synapsen (synaptische Plastizität) werden als biologische Grundlage des Gedächtnisses angesehen.
Für die Funktion des Gehirns scheint insgesamt also weniger die Anzahl der Nervenzellen als die Zahl deren Verknüpfungen wesentlich. Die Verminderung der Anzahl der Nervenzellen scheint manchmal sogar Bedingung für deren gute Funktion. Dabei ist das Wechselspiel zwischen Erregung in einer Hirnregion und gleichzeitiger Hemmung in einer anderen Region entscheidend. Für eine gute reibungslose Funktion des Gehirns ist Bedingung, dass bei es bei jeder Aktivierung eine Hemmung an anderer Stelle gibt.
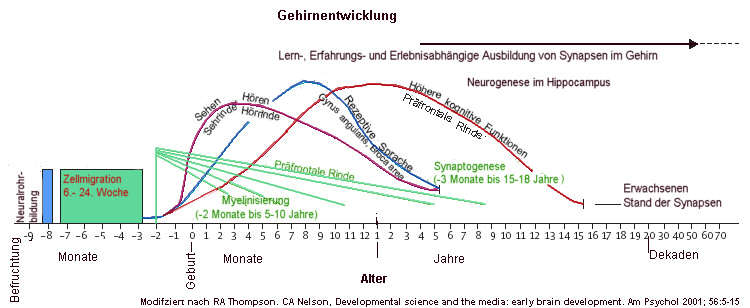
Die Verknüpfungen der Nervenzellen (Synapsen) ist entscheidend für die Speicherung komplexer Informationen und Gefühle. Wenn Synapsen besonders häufig benutzt werden, wird deren Funktion über verschiedene Mechanismen weiter gebahnt. Nervenzellen verstärken ihre Verbindungen bevorzugt dann, wenn die neuronale Aktivität zwischen ihnen korreliert ist, also gleichzeitig stattfindet. Training und Lernen führen bereits nach kurzer Zeit zu einer kortikalen Reorganisation, die man auch als Nutzungs- oder Erfahrungsabhängige Plastizität bezeichnet.
Synapsen verändern sich also bei Lernvorgängen, die synaptischen Netzwerke der der Nervenzellen werden effektiver, unnötiges kann damit auch entfernt werden. Oft münden die Synapsen zahlreicher präsynaptischer Nervenzellen (selten bis zu 500 000 oft über 10 000) auf ein postsynaptisches Neuron. Jedes Neuron kommuniziert nicht nur mit einem Nachbarn, sondern kann Tausende von Kontakten mit anderen knüpfen. Eine Nervenzelle kann so von einem Nachbarn gehemmt werden, während sie von einem anderen das Signal zur Aktivität bekommt.
Wirkung wiederholter Reize auf die Synapse
Ein wesentlicher Bestandteil synaptischer Plastizität (also der Veränderung von Synapsen bei Lernvorgängen) ist die unterschiedliche Reaktion auf wiederholte Aktivierung:
- Die meisten exzitatorischen (erregenden) Synapsen zeigen eine so genannte paired-pulse facilitation (PPF), dies bedeutet, dass ein zweiter Impuls der in einem engen Zeitfenster stattfindet (40 ms) etwas verstärkt wird.
- Inhibitorische (hemmende) Syynapsen zeigen dagegen eine paired-pulse depression (PPD), hier wird der zweite Impuls im Zeitfenster blockiert.
Oft wiederholte Impulse können zu einem Entstehen von neuen Axonen, Dendriten und neuen Synapsen führen. Diese Effekte sind in verschiedenen Hirnregionen und verschiedenen Arten von Zellen unterschiedlich ausgeprägt.
Das Wachsen neuer Neuriten aus denen später Axone oder Dendriten werden beginnt mit der Aktivierung von Membranrezeptoren durch extrazelluläre Schlüsselreize. Dieses Rezeptoren aktivieren eine intrazelluläre Kaskade, die Veränderungen im Aktinzytoskelet hervorruft, die die dortige Symmetrie verändern. Dann werden durch Regulation der Gentranskription, der Mikrotubuli und der Membrandynamik ausgelöst, die den neuen Neuriten stabilisieren. Hierdurch entsteht die Plastizität des Gehirns und die Möglichkeit zu lernen.
Synaptische Plastizität ist damit die zellbiologische Grundlage von Lernen und Gedächtnis. Lernen wird so zu einer Substanzveränderung. Lernprozesse vermehren oder verringern also die synaptischen Kontakte zwischen den Zellen und lassen auf diese Weise bestimmte Netzwerke im Gehirn entstehen.
Lernen, Gedächtnis, die Sprache und die Entwicklung der Persönlichkeit haben eine ständige Neu-Verknüpfung synaptischer Verbindungen zur Grundlage. Die Funktion des Nervensystem wird durch eine komplexe sich ständig verändernde Architektur neuronaler Netzwerke bestimmt. Diese Komplexität entsteht durch die enorme dreidimensionale Verzweigtheit der einzelnen Neurone in der Entwicklung des Gehirns. Entscheidend ist dabei dass die Neurone zur rechten Zeit in die richtige Richtung wachsen und sich dort mit den richtigen zugehörigen Neuronen über Synapsen verbinden können.
Synapsin und Lernvorgänge
Ein wichtiger Mechanismus der Signalübermittlung an der Synapse ist die Proteinphosphorylisation. An den synaptischen Vesikeln (Bläschen) finden sich die drei Phosphoproteine die Synapsin I, Synapsin II, und Synaptophysin. Dephosphoryliertes Synapsin I hält die Neurotransmittervesikel in einem Reservepool, wenn Synapsin I phosphoryliert wird, verschwindet diese Speicherfunktion und die synaptischen Vesikel werden bereit für die Ausschüttung der Neurotransmitter. Diese Regulierung der Transmitterausschüttung durch Phosphorylierung der Synapsine ist ein Mechanismus, über den Lernvorgänge stattfinden. Synapsine sind außerdem auch an der Bildung neuer Synapsen beteiligt.
Studien zeigen, dass ein Fehlen von Synapsin I zu Lernschwierigkeiten führt und die Neuentstehung und Veränderung von Synapsen beeinträchtig (Rosahl et al. 1993). Umgekehrt konnte gezeigt werden, dass das Lösen von Aufgaben, die Lernprozesse auslösen, zu einer vermehrten Expression von Synapsin I führt. Dies insbesondere im für das Gedächtnis wichtigen Hippocampus (Gómez-Pinilla et al 2001).
DARPP-32 und Lernvorgänge
Das Phosphoprotein DARPP-32 findet sich vor allem im Neostriatum, einer Hirnregion die Teil der so genannten Basalganglien ist. DARPP-32 spielt eine große Rolle bei der Integration vieler dort eintreffender Neurotransmittersignale. Bestimmte Neurotransmittersignale führen zu einer Phosphorylisierung und andere bei der Dephosphorylisierung von DARPP-32. Die phosphorylisierte Form, aber nicht die dephosphorylisierte Form, von DARPP-32 hemmt eine Proteinphosphatase, die wiederum verschiedene Ionenkanäle und -pumpen kontrolliert. Die Wirkung vieler Neurotransmitter wird über diese komplexe Signalkaskade gesteuert. Auch hierüber werden Lernvorgänge im Gehirn vermittelt. DARPP-32 beeinflusst dabei sowohl das motorische Lernen (Erlernen von Bewegungsabläufen), als auch das kognitive Lernen (Aneignen von Wissen) (Graybiel 2005).
Genetische Varianten von DARPP-32 können offenbar zu einer besseren Lernfähigkeit führen. Andere Mutationen im DARPP-32-Gen hingegen können Krankheiten wie Parkinson und Schizophrenie begünstigen (Meyer-Lindenberg et al. 2007).
Konzept der Kurzzeit- und Langzeitpotenzierung
Ein Modell für einen zellulären Lernmechanismus ist die Langzeitpotenzierung (LTP) im Gegensatz zur Kurzzeitpotenzierung (STP).
- Vorübergehende synaptische Plastizität nennt man Kurzzeitpotenzierung (STP) (short-term potentiation) oder das Gegenteil short-term depression, STD). Sie spielt bei der Aufmerksamkeitslenkung, dem Arbeitsgedächtnis und der Kontexterfassung eine Rolle.
- Bei der Langzeitpotenzierung (LTP) (long-term potentiation) geht es um die Konsolidierung also um dauerhafte Lernprozesse. LTP scheint die Basis für das fast unerschöpfliche Reservoir unseres Gedächtnis und vieler anderer Vorgänge im Gehirn zu sein.
LTP passiert an allen erregenden Synapsen, die meiste Forschung dazu konzentriert sich aber auf den Hypocampus und die CA1-Synapsen die überwiegend mit dem Neurotransmitter Glutamat arbeiten. Unzweifelhaft findet der Vorgang aber auch in der Hirnrinde statt. Möglicherweise findet die LTP in der Rinde und im Hippocampus aber mit unterschiedlichen Mechaninsmen statt. Im Hippocampus scheint die Zunahme der synaptischen Feuerung eine größere Rolle zu spielen, als der Umbau der Synapsen, in der Rinde umgekehrt.
Eine Langzeitpotenzierung kommt durch die langanhaltende Bahnung synaptischer Übertragung nach einer Aktivierung der Synapse durch intensive hochfrequente Stimulation des präsynaptischen Neurons zustande. Experimente mit Stimulationen durch hochfrequente Stromimpulse im Bereich des Hippocampus über mehrere Sekunden zeigten, dass selbst nach einigen Tagen die Reizung durch Stromimpulse niedriger Intensität noch gebahnt war.
LTP wird über repetitive (wiederholte) Aktivierung von Glutamat-Rezeptoren getriggert. Insbesondere spielen dabei zwei Untergruppen der Glutamat-Rezeptoren eine wichtige Rolle:
- NMDA-Rezeptoren (N-methyl-D-aspartate-Rezeptoren) und
- Ampa-Rezeptoren (A-amino-3-hydroxy-5-methyl-4-isoxazolepropionic-Rezeptoren)
Diese nehmen Einfluss auf die Durchlässigkeit der Na und K- Kanäle und erzeugen einen Einwärtsstrom, der das Ruhemembranpotential verändert. Während der LTP wird vermutlich der AMPA-Rezeptor in seiner Lokalisation verändert, auch die Zahl der Rezeptoren scheint zuzunehmen.
Die Kurzzeitplastizität führt nicht zu strukturellen Veränderungen. Bei der LTP hingegen kommt es zu Veränderungen der Proteinsynthese, zu einem synaptischen Remodelling und zu infrastrukturellen Veränderungen in Zellprozessen. Bei der LTP wird durch einen von außen zugeführten Reiz die Nervenzellantwort dauerhaft verändert, entsprechend einem Lernerfolg.
Die Tatsache, daß LTP und LTD durch korrelierte Aktivität prä- und postsynaptischer Nervenzellen induziert werden, sollte für die Spezifität synaptischer Veränderungen sorgen. Das heißt dafür, dass lediglich solche Synapsen verstärkt werden, die während des Induktionsprozesses aktiv waren.
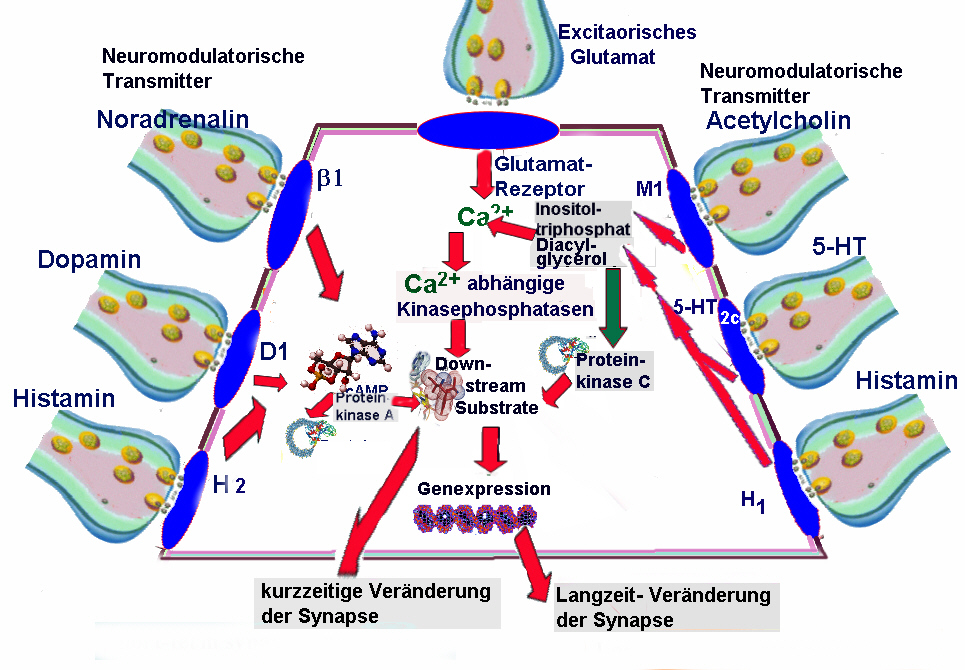
Untersuchungen zeigen allerdings, daß sich nach aktivitätsabhängiger Modifikation einer Gruppe von Synapsen die Übertragungseigenschaften in benachbarten Synapsen ebenfalls verändern. Die wahrscheinlichste Erklärung für eine solche unspezifische Ausbreitung synaptischer Veränderungen sind diffundierende Botenstoffe, die am Ort der Induktion freigesetzt werden und dann auch Synapsen in der unmittelbaren Nachbarschaft beeinflussen (Polnau et al. 2002).
Die Gene, die die LPT und LPD regulieren sind Vitamin A-abhängig. Vitamin A spielt damit bei der Plastizität des Gehirns eine große Rolle. In unseren Breiten dürfte das eine geringe Rolle spielen, da die generelle Versorgung sehr gut ist, in Entwicklungsländern könnte dies mit ein Grund für geistige Entwicklungsverzögerungen sein (Misner et al. 2001).
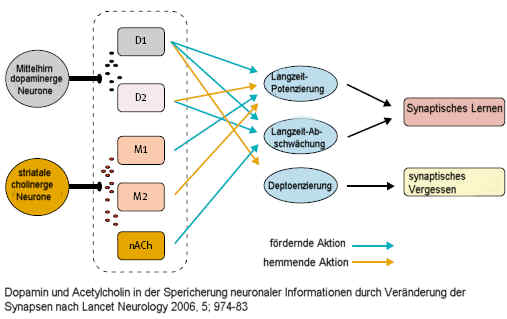
LTP wird durch Schlafmangel behindert, was Schwierigkeiten bei der Konsolidierung von Gedächtnisinhalten bei Schlafmangel erklärt. Körperliche Aktivität hingegen fördert die LTP, die hippocampale Neurogenese, die synaptische Plastizität und damit auch die Gedächtnisbildung und Lernvorgänge.
Lernvorgänge verändern die Genexpression
Erfahrungen und Erlebnisse verändern nicht nur die Anzahl und den Ort der Verbindungsstellen zwischen Nervenzellen, sie verändern auch die Genaktivität der Zelle, d.h. sie beeinflussen, welche Gene wie stark exprimiert/abgelesen werden. Auch über diesen Mechanismus werden Erfahrungen zu bleibenden Teilen der Person.
Lernvorgänge führen zum „Anschalten“ von Genen, die sonst nicht genutzt würden. Eine multimodale Stimulation verstärkt damit neuronale Netze, vorausgesetzt sie verursacht nicht zuviel Stress und überfordert nicht. Eine interessante Umwelt und die Gesellschaft von Menschen fördert damit nicht nur die Verschaltung der Synapsen, sondern verändert auch die Genaktivität der einzelnen Nervenzellen.
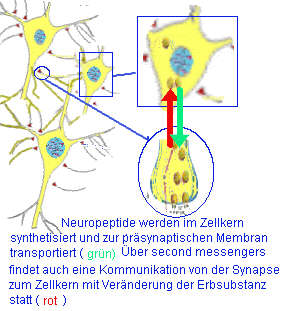
Resultat ist eine große Zahl verschiedener Neuropeptide (> 40), deren Verteilung im Gehirn sich ständig nach den Lernerfahrungen und Anforderungen ändert. Auch Neuropeptide sind chemische Botenstoffe die Informationen im Nervensystem vermitteln.
Neuropeptide findet man in besonders großer Zahl im Hypothalamus, der Hypophyse, den endokrinen Drüsen und im Verdauungssystem. Peptidtransmitter sind beispielsweise Enkephalin, CGRP, VIP und Cholecystokinin.
Herstellung und Transport der Neurotransmitter
In der präsynaptischen Axonendigung befinden sich je nach Zelle wenige (5-10) bis einige Tausend membranumhüllte synaptische Vesikel, die einen Durchmesser von ungefähr 40nm haben und jeweils 1000 bis 5000 Transmittermoleküle enthalten. Wo die Neurotransmitter in der Zelle hergestellt werden, hängt von der Art des Transmitters ab: Die monoaminen Neurotransmitter werden überwiegend direkt im Endknöpfchen hergestellt, die Peptide an den Ribosomen im Soma des Neurons und wandern dann zum Endknöpfchen. Vorstufen mancher Transmittermoleküle aber auch diese selbst werden im Zellkörper hergestellt, sie werden dann über oft lange Strecken (Man bedenke z.B. die Länge des Ischiasnerven vom Rückenmark zum Fuß), durch das Axoplasma, bis zum Ende der Nervenfaser transportiert. Dieser Weg ist langsam (40 cm/Tag) spielt aber nur für die „Vorratshaltung“ und bleibende Veränderungen eine Rolle. Der Transport geht dabei in beide Richtungen, antegrad und retrograd.
„Reuptake“ der Neurotransmitter
Was mit den Neurotransmittern passiert, nachdem sie in den synaptischen Spalt ausgeschüttet haben und ihre Aufgabe der Signalübertragung abgeschlossen ist, ist je nach Transmitter und Synapse unterschiedlich. Während der Transmitter Acetylcholin typischerweise innerhalb des synaptischen Spalts vom Enzym Acetylcholinesterase gespalten und damit inaktiviert wird, werden andere Transmitter wie Serotonin und Dopamin zumindest teilweise wieder in die Präsynapse aufgenommen. Man spricht dabei von „Reuptake“ oder „Wiederaufnahme“.
Diese Wiederaufnahme in die Nervenzelle, die den Transmitter ausgeschüttet hat, ist besonders über die als Antidepressiva eingesetzten Serotonin Wiederaufnahmehemmer (Serotonin-Reuptake-Inhibitoren, SRI) allgemein bekannt geworden. Diese erhöhen durch die Hemmung des Reuptakes die Konzentration des Transmitters in der Synapse und verstärken so die Signalübertragung über die Synapse. Dieser Wirkmechanismus gilt allerdings auch für andere Neurotransmitter und auch andere Antidepressiva.
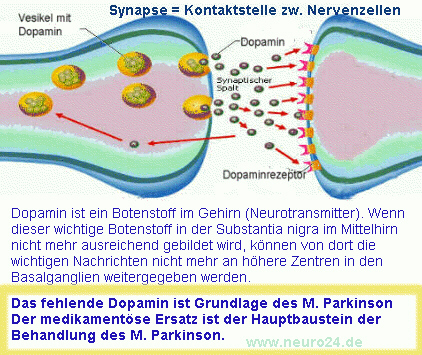
Die Wiederaufnahme der Neurotransmitter spielt auch biologisch bei Persönlichkeitsstörungen und anderen Erkrankungen eine Rolle. Beispiel: Der Dopamintransporter (DAT) vermittelt die synaptische Wiederaufnahme von Dopamin in das dopaminerge Neuron über einen Na+– und CI–-gekoppelten Mechanismus. Der DAT kommt nur in dopaminergen Neuronen vor. Besonders stark exprimiert wird er in den Teilen des Gehirns, die beim Morbus Parkinson besonders von der Neurodegeneration betroffen sind (im ventralen Teil der Substantia nigra pars compacta, weniger in der ventralen Tegmentumregion und dem Hypothalamus). Zunutze machen kann man sich das zum Beispiel beim so genannten DaTSCAN, einer nuklearmedizinischen Untersuchung bei der diese Dopamintransporter sichtbar gemacht werden können und so die Funktionsfähigkeit der dopaminergen Nervenzellen überprüft werden kann.
Wirkung von Medikamenten an der Synapse
Synapsen bilden einen Angriffspunkt für verschiedene Medikamente aber auch für Gifte.
Störbar oder verbesserbar ist die Übertragung bei:
- der Synthese des Transmitters
- der Speicherung des Transmitters
- der Freisetzung des Transmitter
- der Reaktion des Transmitters mit „seinem“ Rezeptor
- der Wiederaufnahme des Transmitters („Reuptake“, siehe oben)
- auf Ebene der ‚Second messenger‚.
Medikamente wie Gifte haben dabei verschiedene Möglichkeiten der Beeinflussung: die Vesikelentleerung kann gehemmt werden oder eine vollständige Entleerung ausgelöst werden. Hemmung der Wiederaufnahme der Spaltprodukte ins Endknöpfchen ( Wiederaufnahme- oder Reuptake-Hemmer), Hemmung der Resynthese oder Speicherung, Hemmung des Transmitterabbaus, Blockierung der Rezeptoren durch „falsche“ Transmitter, Agonisten können den Effekt des Transmitters imitieren, Antagonisten blockieren die Bindung und öffnen die Ionenkanäle nicht.
Selbstverständlich haben die Neurotransmitter nicht nur erregende Funktion, sondern genau so wichtig auch hemmende. Hemmende und erregende Synapsen bewirken eine Verrechnung durch räumliche und zeitliche Summation. Nur wenn das Ergebnis der Verrechnung aller Synapsen eine überschwellige Depolarisierung am Axonhügel des Zellkörpers bewirkt, leitet das Axon eine Erregung weiter. Eine Störung der Balance zwischen Erregung und Inhibition ist die Ursache vieler neurologischer Erkrankungen von M. Parkinson bis Epilepsie. Die medikamentöse Behandlung beruht hier weitgehend auf einem Konzept, das auf Interaktionen von chemischen Synapsen aufbaut.
Die Wirkung von Medikamenten, die die Signalübertragung an Synapsen beeinflussen, hängt auch davon ab, wie schnell die Veränderungen an der Synapse eintreten bzw. wie schnell die Substanz an der Synapse ankommt. Codein muss erst in der Leber zu Morphium umgewandelt werden und kommt deshalb erst langsam als Morphium zum Gehirn, es euphorisiert deshalb meist nicht. Das chemisch vom Morphium nur gering abgewandelte Heroin (Diacetylmorphin) hingegen ist fettlöslicher als Morphium und kann deshalb viel schneller als Morphium vom Gehirn aufgenommen werden. Hierdurch kommt es zur Euphorisierung und schnellen Suchtentwicklung.
Kann Psychotherapie wie ein Medikament wirken?
Wenn Menschen miteinander sprechen, kommuniziert das eine Gehirn mit dem anderen, erzeugt dort durch Lernprozesse anatomische Veränderungen und umgekehrt. Es verwundert deshalb nicht, dass funktionelle Kernspinaufnahmen bei Wirkung einer Psychotherapie teils ähnliche Veränderungen im Gehirn und damit auch an den Synapsen zeigen wie dies bei Behandlung mit Psychopharmaka der Fall ist.
